РЕПЛИКАЦИЯ (от позднелат. replicatio - повторение) (редупликация), самовоспроизведение нуклеиновых кислот (обычно ДНК, у некоторых вирусов РНК), обеспечивающее точное копирование генетич. информации и передачу ее от поколения к поколению. При Р. ДНК нуклеотидная последовательность копируется (целиком или частично) в виде комплементарной последовательности (см. Комплементарность) дезоксирибонуклеотидов.
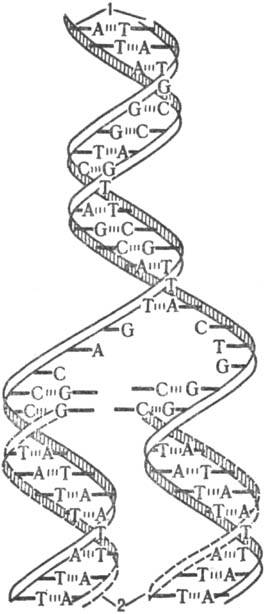
В процессе Р. двойная спираль ДНК, состоящая из двух комплементарных полинуклеотидных цепей, раскручивается на отдельные цепи и одновременно начинается синтез новых полинуклеотидных цепей; при этом исходные цепи ДНК играют роль матриц. Новая цепь, синтезирующаяся на каждой из исходных цепей, идентична др. исходной цепи. Когда процесс завершается, образуются две идентичные двойные спирали, каждая из которых состоит из одной старой (исходной) и одной новой цепи (рис. 1). Таким образом от одного поколения к другому передается только одна из двух цепей, составляющих исходную молекулу ДНК,-т. наз. полуконсервативный механизм Р.
Р. состоит из большого числа последоват. этапов, которые включают узнавание точки началу Р., расплетание исходного дуплекса (спирали), удержание его цепей в изолированном друг от друга состоянии, инициацию синтеза на них новых дочерних цепей, их рост (элонгацию), закручивание цепей в спираль и терминацию (окончание) синтеза. Все эти этапы Р., протекающие с высокой скоростью и исключит. точностью, обеспечивает комплекс, состоящий более чем из 20 ферментов и белков,-т. наз. ДНК-репликазная система, или реплисома. Функцион. единица Р.-реплик он, представляющий собой сегмент (участок) хромосомы или внехромосомной ДНК, ограниченный точкой начала, в которой инициируется Р., и точкой окончания, в которой Р. останавливается. Скорость Р. контролируется на стадии инициации. Однажды начавшись, Р. продолжается до тех пор, пока весь репликон не будет дуплицирован (удвоен). Частотд инициации определяется взаимод. спец. регуляторных белков с точкой начала Р. Бактериальные хромосомы содержат один репликон: инициации в единств. точке начала Р. ведет к Р. всего генома. В каждом клеточном цикле Р. инициируется только один раз, Плазмиды и вирусы, являющиеся автономными генетич. элементами, представляют собой отдельные репликоны, способные к многократной инициации в клетке-хозяине. Эукариотич. хромосомы (хромосомы всех организмов, за исключением бактерий и синезеленых водорослей) содержат большое число репликонов, каждый из которых также однократно инициируется за один клеточный цикл.
Начиная с точки инициации, Р. осуществляется в ограни-ченной зоне, перемещающейся вдоль исходной спирали ДНК. Эта активная зона Р. (т. наз. репликац. вилка) может двигаться в обоих направлениях. При однонаправленной Р. вдоль ДНК движется одна репликац. вилка. При Двунаправленной Р. от точки инициации в противоположных направлениях расходятся две репликац. вилки; скорости их движения могут различаться. При Р. ДНК бактерии и млекопитающих скорость роста дочерней цепи составляет соотв. 500 и 50 нуклеотидов в 1 с; у растений эта величина не превышает 20 нуклеотидов в 1 с. Движение двух вилок в противоположных направлениях создает петлю, которая имеет вид "пузыря" или "глаза". Продолжающаяся Р. расширяет "глаз" до тех пор, пока он не включит в себя весь репликон.
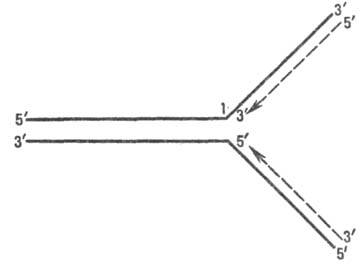
В ходе Р. рост цепи осуществляется благодаря взаимод. дезоксирибонуклеозидтрифосфата с 3'-ОН концевым ну-клеотидом уже построенной части ДНК; при этом отщепляется пирофосфат и образуется фосфодиэфирная связь. Рост полинуклеотидной цепи (рис. 2) идет только с ее З'-конца, т. е. в направлении 5' : 3' (см. Нуклеиновые кисло-ты). Фермент, катализирующий эту реакцию,-ДНК-полиме-раза (см. Полидезоксирибонуклеотид-синтетазы)-не способен начать матричный синтез на одноцепочечной ДНК, если нет хотя бы олигонуклеотидного биспирального участка (т. наз. затравочного олигонуклеотида) комплементарного матрице; затравочным олигонуклеотидом во мн. случаях является не ДНК, а РНК.
Рис. 2. Направление роста дезоксирибонуклеотидных цепей при репликации; сплошные линии - исходная ДНК, пунктирные - новые цепи ДНК (стрелки показывают направлениеих роста); 1-репликац. вилка.
Энергия, затрачиваемая на образование каждой новой фосфодиэфирной связи в цепи ДНК, обеспечивается расщеплением фосфатной связи между a- и b-фосфатными группами нуклеозидтрифосфата.
ДНК-полимераза имеет один центр связывания нуклеозидтрифосфата, общий для всех четырех нуклеотидов. Выбор из среды нуклеотида, основание которого комплементарно очередному основанию матрицы, протекает без ошибок, благодаря определяющему влиянию ДНК-матрицы (исходной цепи ДНК). При некоторых мутационных повреждениях структуры ДНК-полимеразы в ряде случаев происходит включение некомплементарных нуклеотидов.
В процессе Р. формальной ДНК на короткое время с вероятностью 10-4-10-5 возникают редкие таутомерные формы всех 4 азотистых оснований нуклеотидов, которые образуют неправильные пары. Высокая точность Р. (вероятность ошибок не превышает 10-9) обусловлена наличием механизмов, осуществляющих коррекцию (репарацию).
Репликац. вилка асимметрична. Из двух синтезируемых дочерних цепей ДНК одна строится непрерывно, а другая-с перерывами. Первую наз. ведущей, или лидирующей, цепью, а вторую-отстающей. Синтез второй цепи идет медленнее; хотя в целом эта цепь строится в направлении 3' : 5', каждый из ее фрагментов в отдельности наращивается в направлении 5' : 3' (рис. 3). Благодаря такому прерывистому механизму синтеза, Р. обеих антипараллельных цепей осуществляется с участием одного фермента-ДНК-полимеразы, катализирующего наращивание нуклеотидной цепи только в направлении 5' : 3'.
Рис. 3. Схема механизма роста цепей ДНК при репликации: А-ведущая цепь, Б-отстающая цепь, В-фрагмент Оказаки.
В качестве затравок для синтеза фрагментов отстающей цепи служат короткие отрезки РНК, комплементарные матричной цепи ДНК. Эти РНК-затравки (праймеры), состоящие примерно из 10 нуклеотидов, с определенными интервалами синтезируются на матрице отстающей цепи из рибонуклеозидтрифосфатов в направлении 5' : 3' с помощью фермента РНК-праймазы. РНК-праймеры затем наращиваются дезоксинуклеотидами с 3'-конца ДНК-поли-меразой, которая продолжает наращивание до тех пор, пока строящаяся цепь не достигает РНК-затравки, присоединенной к 5'-концу предыдущего фрагмента. Образующиеся таким образом фрагменты (т. наз. фрагменты Оказаки) отстающей цепи насчитывают у бактерий 1000-2000 дез-оксирибонуклеотидных остатков; в животных клетках их длина не превышает 200 нуклеотидов.
Чтобы обеспечить образование непрерывной цепи ДНК из многих таких фрагментов, в действие вступает особая система репарации ДНК, удаляющая РНК-затравку и заменяющая ее на ДНК. У бактерий РНК-затравка удаляется нуклеотид за нуклеотидом благодаря 5' : 3'-экзонуклеазной активности ДНК-полимеразы. При этом каждый отщепленный рибонуклеотидный мономер замещается соответствующим дезоксирибонуклеотидом (в качестве затравки используется З'-конец синтезированного на старой цепи фрагмента). Завершает весь процесс фермент ДНК-лигаза, катализирующий образование фосфодиэфирной связи между группой З'-ОН нового фрагмента ДНК и 5'-фосфатной группой предыдущего фрагмента. Образование этой связи требует затраты энергии, которая поставляется в ходе сопряженного гидролиза пирофосфатной связи кофермента-никотинамид-адениндинуклеотида (в бактериальных клетках) или АТФ (в животных клетках и у бактериофагов).
Раскручивание двойной спирали и пространств. разделение цепей осуществляется при помощи неск. спец. белков. Т. наз. геликазы расплетают короткие участки ДНК, находящиеся непосредственно перед репликац. вилкой. На разделение каждой пары оснований расходуется энергия гидролиза двух молекул АТФ до аденозиндифосфата и фосфата. К каждой из разделившихся цепей присоединяется неск. молекул ДНК-связывающих белков, которые препятствуют образованию комплементарных пар и обратному воссоединению цепей. Благодаря этому нуклеотидные последовательности цепей ДНК оказываются доступными для репликативной системы. Др. специфич. белки помогают праймазе получить доступ к матрице отстающей цепи. В результате праймаза связывается с ДНК и синтезирует РНК-затравки для фрагментов отстающей цепи. Для формирования новых спира-,лей не требуется ни затрат энергии, ни участия к.-л. "закручивающего" фермента.
В случае кольцевого репликона (напр., у плазмиды) описанный процесс наз. q-репликацией. Т.к. кольцевые молекулы ДНК закручены сами на себя (суперспирализо-ваны), при раскручивании двойной спирали в процессе Р. они должны непрерывно вращаться вокруг собств. оси. При этом возникает торсионное напряжение, которое устраняется путем разрыва одной из цепей. Затем оба конца сразу же вновь соединяются друг с другом. Эту ф-цию выполняет фермент ДНК-топоизомераза. Р. в этом случае обычно происходит в двух направлениях, т.е. существуют две репликац. вилки (рис. 4). После завершения Р. появляются две двухцепочечные молекулы, которые сначала связаны друг с другом как звенья одной цепи. При их разделении одно из двух колец временно разрывается.
Рис. 4. Один из механизмов репликацииплазмиды (начало репликации обозначено точками); направления движения репликац. вилки показаны стрелками, образующиеся новые цепи ДНК-пунктиром.
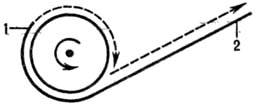
Альтернативный вариант Р. кольцевого репликона предполагает разрыв в одной из цепей двухспиральной молекулы ДНК. Образовавшийся при этом свободный 3'-конец кова-лентно наращивается, оставаясь связанным с матрицей (второй, неразорванной цепью), а 5'-конец постепенно вытесняется новой полинуклеотидной цепью (рис. 5). Таким образом одна цепь разматывается и непрерывно удлиняется, а репликац. вилка скользит вокруг кольцевой матричной цепи (механизм "катящегося кольца"). По мере роста новой цепи вытесненная цепь с освободившимся 5'-концом становится линейной матрицей для синтеза новой комплементарной цепи. Этот синтез на линейной матрице продолжается до тех пор, пока не образуется дочерняя цепь ДНК, комплементарная одному обороту кольцевой матрицы, т. е. целому репликону. Таким путем с кольцевой матрицы может сходить большое число комплементарных копий. Такой механизм обнаружен у некоторых вирусов, а также в ряде клеток эукариот.
Рис. 5. Схема репликации по механизму катящегося кольца (новая молекула ДНК показана пунктиром): 1-3'-конец ДНК; 2-5'-конец ДНК.
Еще одна схема Р. предполагает формирование структуры, названной D-петлей. Согласно этому механизму, сначала реплицируется только одна из цепей кольцевого репликона, тогда как вторая цепь, оставаясь интактной, вытесняется, образуя петлю. Р. второй цепи начинается с др. стартовой точки и только после того, как реплицировалась часть первой цепи. Такой механизм Р. обнаружен, например, у митохондриальных ДНК.
Р. РНК (синтез РНК на РНК-матрице) изучена меньше. Она осуществляется только у некоторых вирусов (напр., у вирусов полиомиелита и бешенства). Фермент, катализирующий этот процесс,-РНК-зависимая РНК-полимераза (его называют также РНК-репликазой или РНК-синтетазой). Известно неск. типов Р, РНК: 1) вирусы, содержащие матричные РНК, или мРНК [т. наз. (+)РНК], в результате Р. образуют комплементарную ей цепь [(-)РНК], не являющуюся мРНК, которая используется как матрица для синтеза (+)РНК; 2) вирусы, содержащие (—)РНК, в результате Р. синтезируют (+)РНК; 3) вирусы, содержащие двухцепочечную РНК [(+)PHK и (—)РНК], в результате асимметрической Р. синтезируют (+)РНК.
Гипотеза о механизме Р. сформулирована в 1953 Дж. Уотсоном и Ф. Криком, которые предположили, что две комплементарные цепи ДНК после их разделения могут выполнять ф-ции матриц для образования на них новых цепей ДНК. В 1958 М. Мезельсон и Ф. Сталь экспериментально подтвердили такой механизм Р.
Лит.: Степт Г., Кэлиндар Р., Молекулярная генетика, пер. с англ., М., 1981, с. 499-520; Kornberg А., DNA replication, S. F., 1980; Ogawa T, Oka-zaki Т., "Ann. Rev. Biochem.", 1980, v. 49, p. 421-57. П.Л. Иванов.