МАТРИЧНЫЕ РИБОНУКЛЕИНОВЫЕ КИСЛОТЫ (информационные РНК, мРНК, иРНК), молекулы РНК, представляющие собой комплементарные (см. Комплементарность) копии участков значащих цепей генов (ДНК), в которых закодирована информация об аминокислотных последовательностях полипептидных цепей белков. Образуются в результате транскрипции (синтез РНК на ДНК-матрице с помощью фермента ДНК-зависимой РНК-полимеразы) и послед. процессинга первичного транскрипта - удаления избыточных РНК-последовательностей с обоих концов молекулы и некодирующих последовательностей РНК, а также сшивания кодирующих участков РНК друг с другом. мРНК переносят генетич. информацию от генов к рибосомам, где происходит трансляция (синтез полипептидных
цепей на мРНК-матрице). При этом одну цепь мРНК могут одновременно транслировать неск. рибосом (т. наз. полирибосомы, или полисомы).
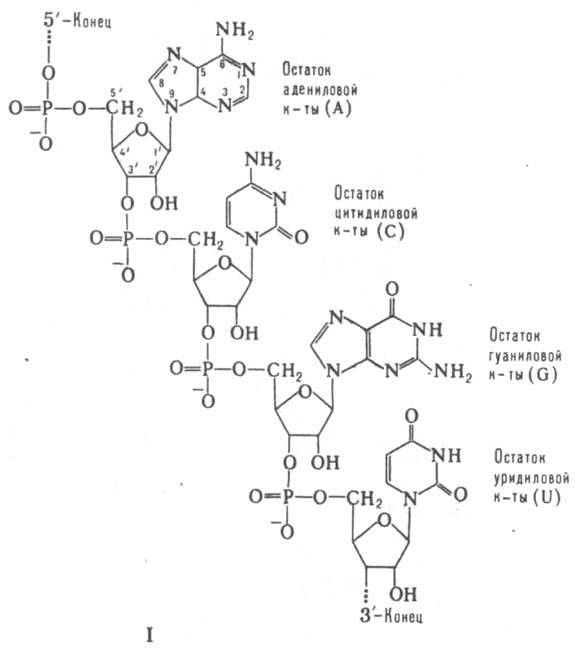
мРНК всегда состоят из одной полинуклеотидной цепи, т. е. относятся к однотяжевым РНК. Как и любые др. РНК, мРНК включают остатки адениловой (А), гуаниловой (G), уридиловой (U) и цитидиловой (С) кислот, связанных между собой 3'-5'-фосфодиэфирными связями (ф-ла I). Вторичная структура мРНК представлена многочисл. двухспиральными участками ("шпильками"), образующимися в результате комплементарного спаривания оснований (А с U и G с С) одной и той же цепи; шпильки соединены между собой короткими однотяжевыми участками. При связывании с рибосомой макромолекулярная структура мРНК существенно изменяется, а двухспиральные ее участки в районе непосредств. контакта с рибосомой переходят в однотяжевое состояние.
мРНК составляют незначит. долю (3-5%) от суммарной клеточной РНК. В клетке бактерий мРНК весьма нестабильны и быстро разрушаются: время их полужизни 2-3 мин. В отличие от них, мРНК в клетках эукариот (все организмы, за исключением бактерий и синезеленых водорослей), как правило, сохраняются в течение неск. часов. В значит. мере это связано с тем, что в эукариотич. клетках мРНК всегда находятся в комплексе с белками, образуя матричные рибонуклеопротеидные частицы, или информосомы. Белковый состав информосом в период, предшествующий трансляции, и в момент трансляции (т.е. в составе полисом) различен.
Молекулы мРНК содержат два типа нуклеотидных последовательностей - транслируемые и нетранслируемые. Транслируемый участок мРНК представляет собой совокупность тринуклеотидов (кодонов; см. Генетический код), кодирующих полипептидную цепь. Он начинается с инициирующего кодона (в подавляющем числе случаев AUG) и заканчивается терминирующим (UAG, UAA или UGA). Одна молекула мРНК прокариот (бактерии и синезеленые водоросли) часто кодирует две или более разных полипептидных цепи (такие мРНК наз. полицистронными) и содержит соотв. неск. кодирующих последовательностей. Аналогичным образом организованы РНК многих РНК-содержащих
бактериофагов (вирусы бактерий), которые в клетках бактерий выполняют ф-ции мРНК.
В полицистронных мРНК последовательности, кодирующие полипептидные цепи, разделены межцистронными нетранслируемыми участками (спейсерами), длина которых варьирует в широких пределах. Между инициирующим кодоном ближайшей к 5'-концу мРНК транслируемой последовательности и этим концом обычно находится 50-200 нуклеотидных остатков. 3'-Конец мРНК отделен от терминирующего кодона ближайшей к нему кодирующей последовательности, как правило, значительно более длинным нетранслируемым участком (до неск. тыс. нуклеотидных остатков). Эти нетранслируемые области, по-видимому, необходимы для создания специфич. макромолекулярной структуры мРНК, а также содержат т. наз. сигналы инициации трансляции. наиб. хорошо изучена такая сигнальная последовательность у мРНК прокариот (открыта Дж. Шайн и Л. Дальгарно и носит их имя), содержащая 3-7 нуклеотидных остатков, расположенных перед инициирующим ко-доном (т. е. ближе к 5'-концу молекулы) каждой транслируемой последовательности. Последовательность Шайн-Дальгарно комплементарна 3'-концевому участку РНК малой субчастицы рибосом и, по всей вероятности, образует с ним в процессе инициации трансляции двухспиральный комплекс.
Вторичная структура мРНК в районе сигнальной последовательности и инициирующего кодона оказывает влияние на эффективность инициации трансляции. Наличие или отсутствие своб. 5'-концевого участка в мРНК прокариот несущественно для начала трансляции. В отдельных случаях рибосома способна начинать трансляцию внутр. кодирующих последовательностей.
Некоторые мРНК прокариот (чаще всего кодирующие ферментыбиосинтеза аминокислот) кроме основных транслируемых последовательностей ближе к их 5'-концу содержат регуляторную последовательность, в которой закодирован короткий полипептид (этот район мРНК носит назв. аттенюатора). Синтез этого полипептида происходит во время образования мРНК и может приводить к преждевременному окончанию синтеза последней.
У некоторых РНК-содержащих вирусов эукариот в РНК закодировано большое число белков, которые в процессе трансляции синтезируются в виде гигантской единой полипептидной цепи (полипротеина), расчленяемой затем специфич. протеазами на отдельные белковые молекулы.
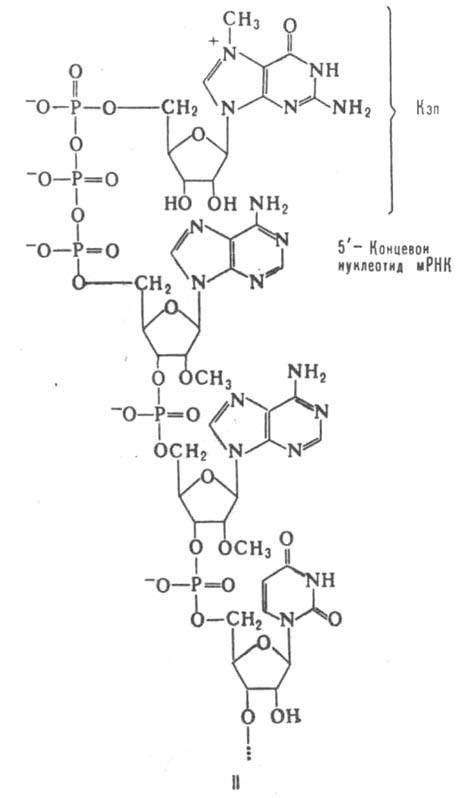
У эукариот мРНК, как правило, моноцистронны. 5'-Конец таких мРНК часто несет модифицир. группировку (т. наз. кэп от англ. cap - головка, ф-ла II), присоединяемую к мРНК после завершения транскрипции с помощью спец. ферментной системы. 5'-Концевые остатки самой полинуклеотидной цепи мРНК, связанные с кэпом, также модифицированы -обычно они содержат остатки рибозы, метилированные по положению 2'. Наличие кэпа в мРНК повышает эффективность ее трансляции. Предполагают, что у эукариот рибосома первоначально связывается с 5'-концом молекулы, а затем "скользит" по цепи мРНК вплоть до инициирующего кодона транслируемой последовательности.
Б. ч. молекул мРНК в эукариотич. клетке содержит на своем 3'-конце гомополимерную цепь (полиаденилат), в которой находится от 20 до 200 остатков адениловой кислоты. Полиаденилирование осуществляется ферментом полиаденилатполимеразой после окончания транскрипции как последний этап процессинга 3'-концевой области мРНК. Функцион. роль полиаденилатной последовательности в мРНК достоверно не установлена; возможно, она служит для связывания спец. белков в информосоме. Некоторые мРНК (напр., гистоновые мРНК) полиаденилированы лишь частично.
Благодаря наличию полиаденилатной последовательности мРНК м. б. легко отделены от остальных клеточных РНК путем избират. комплементарного связывания с
иммобилизованными олигомерами уридиловой или тимидиловой кислоты.
Полиаденилатные последовательности на 3'-концах мРНК часто используют также для получения их ДНК-копий с помощью олигомера тимидиловой кислоты в качестве затравки и фермента обратной транскриптазы. Эти копии в свою очередь используют для получения индивидуальных генов методами генетич. инженерии.
Существование мРНК предсказано в 1956-58 А. С. Спириным и А. Н. Белозерским, а также независимо от них Э. Волкиным и Ф. Астрачаном. Экспериментально мРНК открыты в 1961 в лабораториях М. С. Мезелсона, Дж. Д. Уотсона и С. Спигелмена.
Лит.: Информационные макромолекулы, пер. с англ., М., 1965; Спирин А. С., Молекулярная биология. Структура рибосомы и биосинтезбелка, М., 1986; Льюин Б., Гены, пер. с англ., М., 1987. А. А. Богданов.