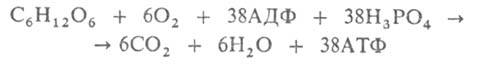Брожение, ферментативное расщепление органических веществ, преимущественно углеводов. Может осуществляться в организме животных, растений и многих микроорганизмов без участия или с участием О2 (соответственно анаэробное или аэробное брожение).
В результате окислительно-восстановительных реакций при брожении освобождается энергия (главным образом в виде АТФ) и образуются соединения, необходимые для жизнедеятельности организма. Некоторые бактерии, микроскопические грибы и простейшие растут, используя только ту энергию, которая освобождается при брожении. Общий промежуточный продукт у многих видов брожения - пировиноградная кислота СН3С(О)СООН, образование которой из углеводов в большинстве случаев протекает таким же путем, как в гликолизе. Некоторые виды брожения, происходящие анаэробно под действием микроорганизмов, имеют важное практическое значение.
Спиртовое брожение осуществляется обычно с помощью дрожжей рода Saccharomyces
и бактерий рода Zimomonas по схеме:
Notice: Undefined variable: iko in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 95
Notice: Undefined variable: goo in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
Notice: Undefined variable: mn in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
где НАД(Ф)Н и НАД(Ф) - соотв. восстановленная или окисленная формы никотинамидадениндинуклеотида или никотинамидадениндинуклеотидфосфата. Первая стадия процесса катализируется ферментом пируватдекарбоксилазой, вторая - алкогольдегидрогеназой. Этот вид брожения используют для промышленного получения этанола, а также в виноделии, пивоварении и при подготовке теста в хлебопекарной промышленности. В присутствии О2 спиртовое брожение замедляется или прекращается и дрожжи получают энергию для жизнедеятельности в результате дыхания.
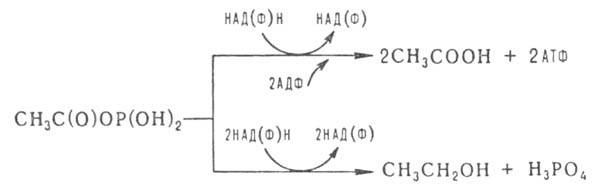
Молочнокислое брожение вызывается бактериями родов Lactobacillus и Streptococcus. При гомоферментативном типе брожение молочная кислота образуется непосредственно из пировиноградной в НАД-зависимой реакции, катализируемой лактатдегидрогеназой. При гетероферментативном брожении метаболизмглюкозы до глицеральдегид-3-фосфата осуществляется по фосфоглюконатному пути по схеме:
Notice: Undefined variable: goo in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
Notice: Undefined variable: mn in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
где АТФ - аденозинтрифосфат, АДФ - аденозиндифосфат. Затем глицеральдегид-3-фосфат
по тому же пути, как в гликолизе. окисляется до молочной кислоты. Образующийся смешанный ангидрид уксусной и фосфорной кислот превращается в уксусную кислоту или восстанавливается до этанола:
Notice: Undefined variable: goo in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
Notice: Undefined variable: mn in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
Молочнокислое брожение играет важную роль при получении различных молочных продуктов (кефира, простокваши и др.), квашении овощей, силосовании кормов в сельском хозяйстве; гомоферментативный процесс используют для промышленного синтеза молочной кислоты. Пропионовокислое брожение идет под действием пропионовокислых бактерий:
Notice: Undefined variable: goo in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
Notice: Undefined variable: mn in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
где SKoA-остаток кофермента A (KoASH), ФАДН и ФАД - соответственно восстановленная и окисленная формы флавинадениндинуклеотида, ~ высокоэргич. связь. Синтез пропионил-КоА катализируется метилмалонил-КоА-карбоксилтрансферазой (кофермент - биотин), а пропионовой кислоты -транстиоэстеразой. Образующийся сукцинил-КоА под действием L-метилмалонил-КоА-мутазы (кофермент - витамин В12) превращается в метилмалонил - КоА, который снова вовлекается в процесс. Параллельно с основными реакциями под действием пируватдегидрогеназы происходит окислительное декарбоксилирование пировиноградной кислоты:
Notice: Undefined variable: goo in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
Notice: Undefined variable: mn in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
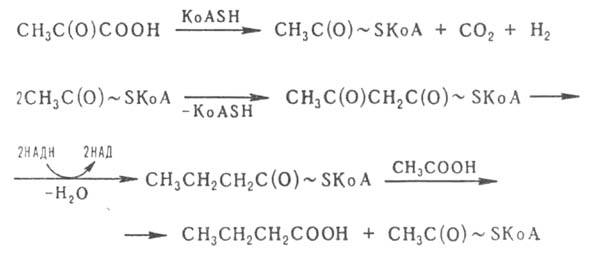
Пропионовокислое брожение используется в молочной промышленности при изготовлении многих твердых сыров. Маслянокислое брожение осуществляется под действием спорообразующих бактерий рода Clostridium по схеме:
Notice: Undefined variable: goo in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
Notice: Undefined variable: mn in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
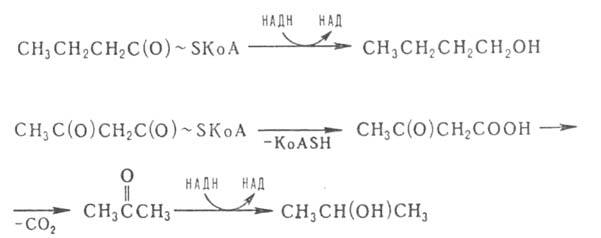
Синтез ацетил-КоА катализируется комплексом ферментов с участием ферродоксина и тиаминдифосфата (тиаминпирофосфата). Из промежуточных продуктов некоторые маслянокислые бактерии синтезируют бутанол, ацетон и изопропанол (так называемое ацетоно-бутиловое брожение):
Notice: Undefined variable: goo in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
Notice: Undefined variable: mn in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
В результате деятельности маслянокислых бактерий Clostridium Kluyreri возможен синтез масляной кислоты из этанола и уксусной или пропионовой кислоты. Механизм реакции связан с окислениемэтанола до СН3С(О) ~ SKoA, который превращается в масляную кислоту. Капроновая кислота образуется в результате взаимодействия бутирил-КоА с ацетил-КоА.
Маслянокислое брожение приводит к порче пищевых продуктов, вспучиванию сыра и банок с консервами. Раньше использовалось для получения масляной кислоты, бутилового спирта и ацетона. Метановое брожение начинается с разложения сложных веществ (например, целлюлозы) до одно- или двууглеродных молекул (СО2, НСООН, СН3СООН и др.), которое осуществляют микроорганизмы, живущие в симбиозе с метанообразующими бактериями. Последние синтезируют метан по схеме:
Notice: Undefined variable: goo in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
Notice: Undefined variable: mn in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
Восстановление СО2 до группы СН3 происходит с участием тетрагидрофолиевой кислоты (ТГФ), затем группа СН3 переносится на витамин В12, где с участием НАДН и АТФ восстанавливается до СН4. В восстановлении др. субстратов ТГФ не участвует.
Метановое брожение в природе происходит в заболоченных водоемах. Используется в промышленных и бытовых очистных сооружениях для обезвреживания органических веществ сточных вод. Образующийся при этом метан (главным образом, в смеси с СО2) используется как топливо.
При аэробном брожение образующийся из пировиноградной кислоты ацетил-КоА конденсируется со щавелевоуксусной кислотой с образованием лимонной, которая претерпевает дальнейшие превращения в цикле трикарбоновых кислот. Суммарное уравнение этого процесса:
Notice: Undefined variable: goo in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
Notice: Undefined variable: mn in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
Продуктами аэробного брожение могут быть метаболиты цикла трикарбоновых кислот:
лимонная, янтарная, фумаровая и другие. В норме они не накапливаются, однако имеются штаммы, главным образом микромицетов, способные накапливать эти соединения в больших количествах. Например, при лимоннокислом брожение выход продукта может достигать 70%, что обусловлено повышенной активностью в микроорганизме цитратсинтетазы.
Интенсивное накопление фумаровой кислоты происходит при функционировании цикла трикарбоновых кислот и глиоксилатного цикла.
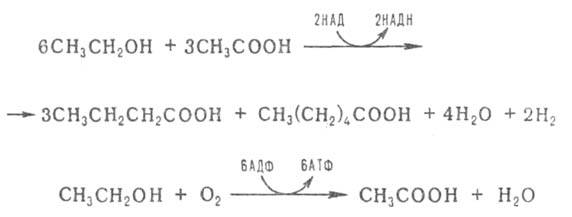
Под действием некоторых аэробных микроорганизмов происходит брожение, при котором
углеродный скелет субстрата не подвергается изменениям. К такому виду брожения, в частности, относится образование уксусной кислоты из этанола (уксуснокислое брожение) под действием уксуснокислых бактерий:
Notice: Undefined variable: goo in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
Notice: Undefined variable: mn in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
Лит.: Кретович В. Л, Биохимия растений, М., 1980, с. 197-224; Брухман Э.Э., Прикладная биохимия, пер. с нем., М., 1981, с. 152-291; Готтшалк Г., Метаболизм бактерий, пер. с англ., М., 1982, с. 186-250; Безбородов A.M., Биохимические основы микробиологического синтеза, М., 1984, с. 173-82.
A.M. Безбородов. Notice: Undefined variable: goo in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109
Notice: Undefined variable: mn in /var/www/chemport.ru/data/www/chemport.ru/chemical_encyclopedia.php on line 109