ХЛОРОФИЛЛЫ (от греч. chloros - зеленый
и phyllon -лист), прир. макрогетероциклич. пигменты, участвующие в процессе
фотосинтеза;
относятся
к металлопорфиринам (см. Порфирины).
Зеленая окраска растений обусловлена присутствием
X., локализованных во внутриклеточных органеллах (хлоропластах или хроматофорах)
в виде пептидных комплексов.
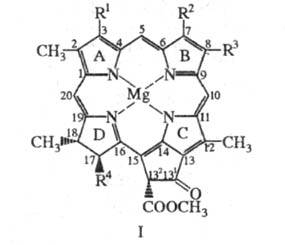
Формально X. представляют собой производные
порфина, молекулы которых содержат циклопентаноновое кольцо, конденсированное
с порфириновым макроциклом, центральный атом Mg и разл. заместители; одно
или два пиррольных цикла в молекулах частично гидрированы, см., например,
ф-лу I. В пиррольном кольце D молекул X. к остатку пропионовой кислоты обычно
присоединены остатки высокомол. изопреноидных спиртов, которые придают X.
способность встраиваться в липидные слои мембран хлоропластов. Для X.,
как и для порфиринов, используется номенклатура ИЮПАК или Фишера.
Из высших растений, водорослей и фотосинтезирующих
бактерий выделено и структурно охарактеризовано св. 50 разл. X. Осн. пигменты
высших растений и зеленых водорослей - X. а
и b. Основа этих
X.- дигидропорфириновый (хлориновый) цикл, содержащий в качестве эфирных
групп (Y) остаток спирта фитола (СН3)2СН(СН2)3СН(СН3)(СН2)3СН(СНз)(СН2)3С(СНз)
= = СНСН2ОН.
При общем содержании X. 0,7-1,1 г на 1
кг зеленой массы растений соотношение X. а и b обычно составляет
3:1 (в зависимости от освещенности, наличия удобрения и др. факторов может
колебаться от 2:1 до 3,4:1, что используется для контроля за развитием
растений). X. а и b выделяют главным образом из листьев крапивы
и шпината (разделяют эти X. хроматографически), X. а - также из
синезеленых микроводорослей, не содержащих X. b.
Близок по структуре к X.
а его
(S)-эпимер по атому С-132 -прир. пигмент X. а', также
участвующий в фотосинтезе. Замена этильной группы в положении 8 в X. а
и
b
на винильную приводит к 8-винилхлорофиллам а
и b,
обнаруженным
в листьях огуречной рассады; участие этих X. в фотосинтезе пока не доказано.
Из бурых и диатомовых водорослей выделены
X. а и с, из красных морских водорослей - X. а
и d.
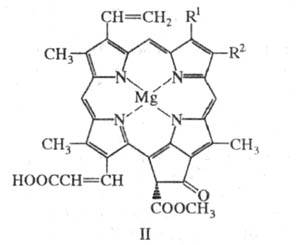
X. группы с (c1,
с2
и c3, ф-ла II) в отличие от др. X. содержат негидрированный
порфириновый макроцикл и остаток неэтерифицированной акриловой кислоты. Находясь
в морских водорослях в виде белковых комплексов, X. этой группы выполняют
в фотосинтезе роль светособирающих антенн.
В большинстве фотосинтезирующих бактерий
обнаружены бактериохлорофиллы (БХ), отличающиеся от X. а типом макроцикла
и замещающими группами в цикле. Они имеют несколько модификаций: так, из
пурпурных бактерий выделены БХ а и b, из зеленых бактерий
- БХ а, с, d и е, из серных бактерий - БХ
с,
d
и
е; обнаружены также фотосинтезирующие бактерии, содержащие БХ g.
В основе БХ a, b
и g
(т.
наз. собственно БХ; ф-ла III) лежит тетрагидропорфириновый макроцикл, содержащий
в качестве эфирных групп (Y) остатки фитола, геранилгераниола (СН3)2С
= СН(СН2)2С(СН3) = СН(СН2)2С(СН3)
= СН(СН2)2С(СН3) = СНСН2ОН
и 2,10-фитадиенола (СНO2СН(СН2)3С(СН3)
= СН(СН2)2СН(СН3)(СН2)3С(СН3)
= СНСН2ОН - для БХ а и b; БХ g содержит
остатки фарнезола
(СН3)2С = СН(СН2)2С(СН3)
= СН(СН2)2С(СН3) = СНСН2ОН
и геранилгераниола. При выделении из ацетона или метанола (особенно в присутствии
оснований) БХ а и b эпимеризуются по атому С-132
с образованием эпимеров БХ а' и b'.
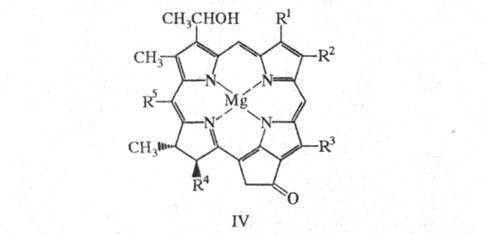
Для БХ с, d и
е (ф-ла
IV), первоначально называемых хлоробиум-хлорофиллами, характерно наличие
дигидропорфиринового макроцикла, a-гидроксиэтильной
группы в положении 3 и разл. алкильных (от С1 до С5)
заместителей в положении 8; эфирные группы (Y) - остатки 2,6-фитадиенола
(СН3)2СН(СН2)3СН(СН3)(СН2)3С(СН3)
= СН(СН2)2С(СН3) = СНСН2ОН
и 2,16,20-фитатриенола (CH3)2C = СН(СН2)2С(CH3)
= СН(СН2)2СН(СH3)(СН2)3
-С(СН3)
= СНСН2ОН.
X.- высокоплавкие интенсивно окрашенные
кристаллы от зеленого до темно-красного и черного цветов; т. пл. X. а
117-121
°С, X. b - 124-125 °С; т. разл. многих X. более 300 °С. X. хорошо
раств. главным образом в полярных орг. растворителях (ДМСО, ДМФА, ацетон, спирты,
диэтиловый эфир), плохо -в петролейном эфире, не раств. в воде. В УФ спектрах
для многих X. характерно наличие
400-430 (т. наз. полоса Соре); полные УФ спектры представлены в табл.
* В скобках даны интенсивности, выраженные
относительно пика в красной области, принятого за единицу.
Под действием кислот и оснований X. легко
подвергаются структурным изменениям. Так, обработка слабыми растворами кислот
приводит к удалению центрального иона Mg с образованием феофитинов. Конц.
НС1 наряду с Mg отщепляет фитол с образованием феофорбидов; раскрытие циклопентанонового
кольца под действием конц. щелочи приводит к хлоринам. При обработке слабыми
щелочами X. постепенно теряют сложноэфирные группы, сохраняя центральный
ионметалла; при этом образуются хлорофиллиды и хлорофиллины. Аналогичные
превращения претерпевают также и бактериохлорофиллы.
Вышеописанные реакции на примере X. а
приведены
на схеме 1.
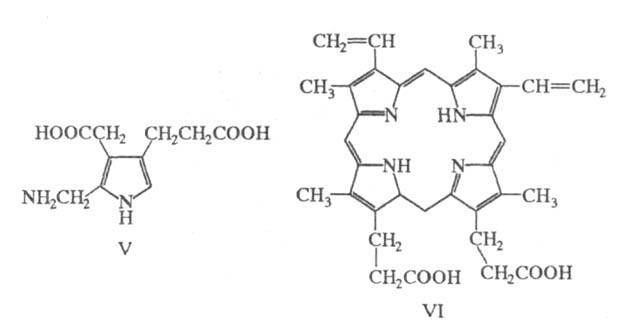
Осн. пути биосинтеза X.: конденсация двух
молекул
аминолевулиновой кислоты с образованием порфобилиногена (ф-ла V), который в
результате ряда ферментативных превращений дает протопорфирин (VI). Из
последнего образуется непосредственный предшественник X.- хлорофиллид,
содержащий атом Mg. Путем последующих реакций восстановления и присоединения
остатков спиртов из него образуется X. Стадия восстановления хлорофиллида
осуществляется у высших растений на свету, у низших - в темноте.
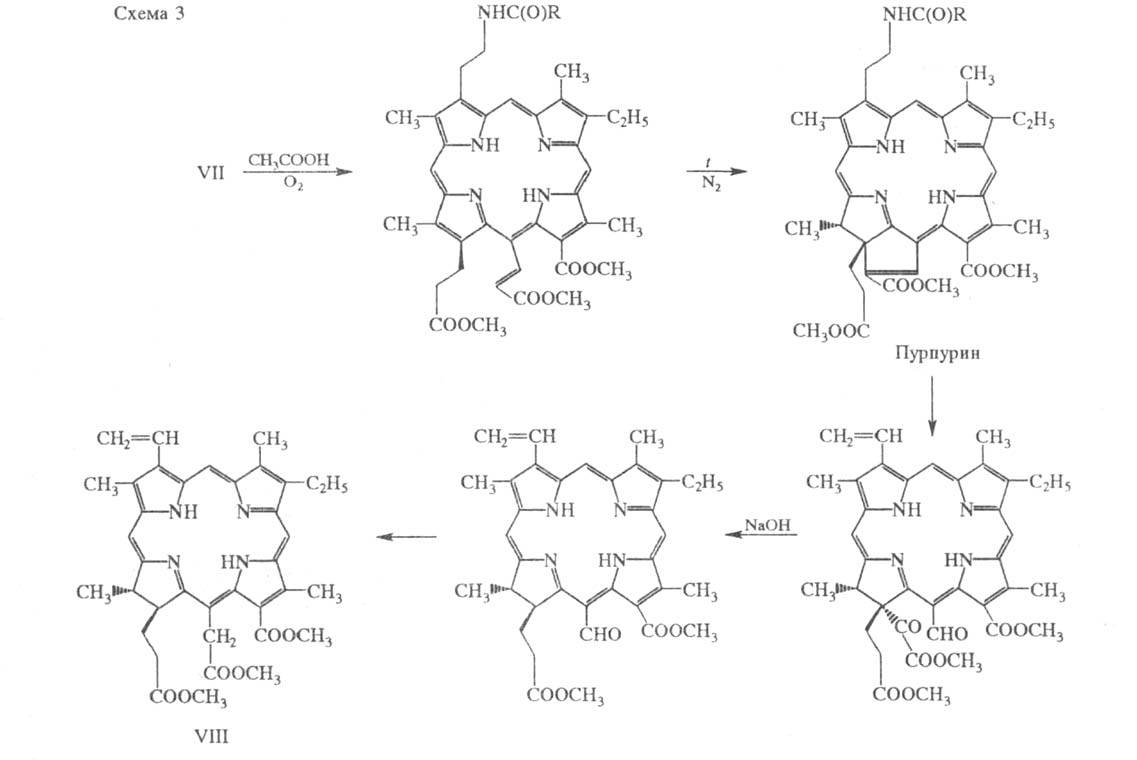
Синтез X. а - одно из замечательных
достижений в области орг. химии. Полный синтез включает 46 стадий. Его
осн. этапы представлены схемами 2, 3. Первый этап заключается в последоват.
синтезе порфирина VII из четырех пирролов -предшественников колец A-D молекулы
X. через два дипир-ролилметана (схема 2). Второй этап - превращение порфирина
VII в триметиловый эфир хлорина е6 (VIII) с транс-конфигурацией
при атомах С-17 и С-18 (т. наз. пурпуриновая реакция; схема 3), послед. циклизация
которого приводит к X. а.
Полный синтез X. а осуществил Р.
Вудворд в 1960.
Лит.: Итоги науки и техники, сер.
Современные проблемы лазерной физики, т. 3, М., 1990; Дайзенхофер И., Михель
X., Фотосинтетический реакционный центр пурпурной бактерии, пер. с нем.,
М., 1990; Woodward R. В., "Pure Appl.Chem.", 1960, №2, p. 383; Woodward
R.B. [а. о.], "Tetrahedron", 1990, v. 46, № 22, p. 7599-7659; Chlorophylls,
ed. by H. Scheer, L., 1991; см. также лит. при ст. Порфирины.

 400-430 (т. наз. полоса Соре); полные УФ спектры представлены в табл.
400-430 (т. наз. полоса Соре); полные УФ спектры представлены в табл.
 нм (
нм (  x 10-3
или отношение интенсивностей к основному пику)
x 10-3
или отношение интенсивностей к основному пику) аминолевулиновой кислоты с образованием порфобилиногена (ф-ла V), который в
результате ряда ферментативных превращений дает протопорфирин (VI). Из
последнего образуется непосредственный предшественник X.- хлорофиллид,
содержащий
аминолевулиновой кислоты с образованием порфобилиногена (ф-ла V), который в
результате ряда ферментативных превращений дает протопорфирин (VI). Из
последнего образуется непосредственный предшественник X.- хлорофиллид,
содержащий